Tidigare forskning har avslöjat att CoV är benägna att överföra arter. Därför, insamlingen av mer information om djur -CoV är avgörande för att förutsäga framtida CoV -utbrott och förhindra zoonotiska överföringshändelser.
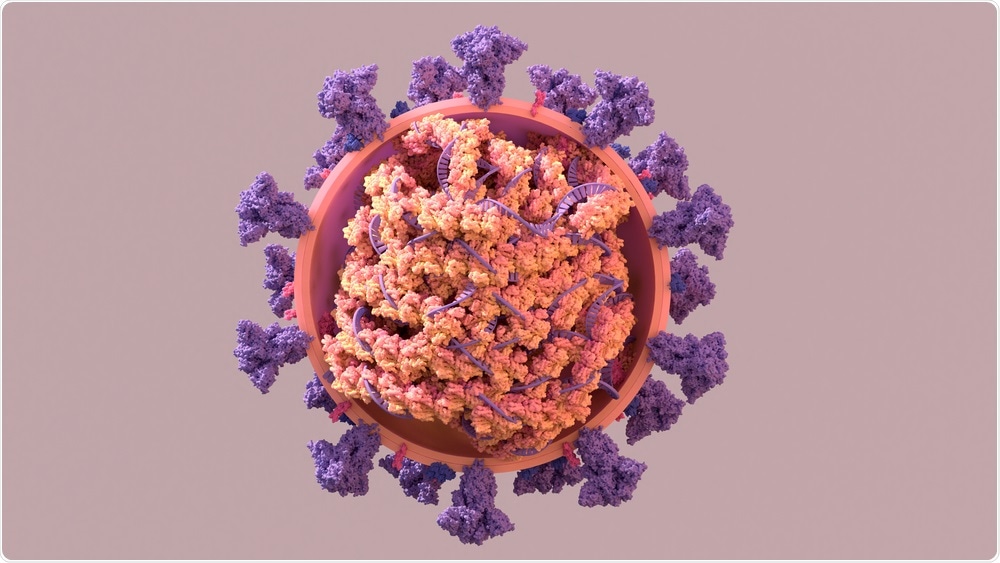 Studie:En jämförande analys av Coronavirus-nukleokapsid (N) -proteiner avslöjar SADS-CoV N-proteinet motverkar IFN-β-produktion genom att inducera ubiquitination av RIG-I. Bildkredit:Design_Cells / Shutterstock.com
Studie:En jämförande analys av Coronavirus-nukleokapsid (N) -proteiner avslöjar SADS-CoV N-proteinet motverkar IFN-β-produktion genom att inducera ubiquitination av RIG-I. Bildkredit:Design_Cells / Shutterstock.com
Forskare har nyligen avslöjat svin akut diarré syndrom (SADS) -CoV, som tillhör släktet Alphacoronavirus, som en ny patogen som orsakar diarré hos nyfödda smågrisar. SADS-CoV, som också kallas svin enteriskt alfakoronavirus (SeACoV), hade rapporterat dödligheten över 35% i södra Kina under ett utbrott 2017.
Förutom SADS-CoV, ytterligare fyra grisar har hittats hittills; nämligen, överförbart gastroenteritvirus (TGEV), svinhemagglutinerande encefalomyelitvirus (PHEV), grisepidemiskt diarrévirus (PEDV), och porcine delta coronavirus (PDCoV). Eftersom SADS-CoV är nära besläktat med bat CoV HKU2-stammar, forskare tror att denna stam har uppstått som ett resultat av genetisk drift eller rekombination förekomster mellan co-infekterande CoV.
Genomiska studier har visat att SADS-CoV innefattar en genetisk sekvens som består av fyra strukturella proteiner, sju oberoende öppna läsramar (ORF) som kodar för sexton icke-strukturella proteiner, och ett tillbehörsprotein, som alla liknar det som finns i många CoV:er. Av de fyra strukturproteinerna nukleokapsid (N) -proteinet innehåller en högkonserverad genomisk sekvens som uttrycks starkt. N -proteinet spelar en roll vid virusinfektion och är också involverat i transkription av subgenomisk ribonukleinsyra (RNA), viral genomreplikation, och dess interaktion med andra proteiner för att stödja virionmontering.
Tidigare studier har föreslagit att SADS-CoV N-proteinet är inblandat i virusets undvikande av värdens medfödda immunsvar, som är kroppens första försvarslinje mot skadliga patogener. Ytterligare, typ I interferon (IFN) signalväg spelar en viktig roll för att skydda värden mot virusinfektion, som inkluderar primär identifiering av patogenassocierade molekylära mönster (PAMP) med mönsterigenkänningsreceptorer (PRR).
Liksom andra RNA -virus, CoV producerar PAMP inklusive dubbelsträngat RNA (dsRNA) och 5′-ppp RNA-intermediärer i cytoplasman under replikation. Dessa PAMP identifieras sedan av värdmönsterigenkänningsreceptorer (PRR), såsom retinsyra-inducerbara gen I (RIG-I) -lika receptorer (RLR). Efterigenkänning och aktivering av RIG-I och/eller melanomdifferentieringsassocierad gen 5 (MDA5) leder till deras interaktion med caspasaktivering och rekryteringsdomäner (CARD).
Senare, prionliknande polymerer bildas, som stimulerar nedströms
TANK -bindande kinas 1 (TBK1) och hämmare av κ B-kinas-ϵ (IKKϵ). Aktiveringen av TBK1 leder till fosforylering av interferon regulatorisk faktor 3 (IRF3) som, i tur och ordning, främjar produktion av typ I IFN. Detta leder i slutändan till uttryck för hundratals IFN-stimulerade gener (ISG).
ISG uttrycks på ett autokrint och parakrint sätt i ett försök att skydda värdcellen från viral invasion. Trots dessa medfödda försvar, virus kan ofta utvecklas för att undvika värdcellens försvar. Till exempel, flera CoV kan hämma värd -IFN -svar under infektion.
En ny studie publicerad i Gränser i immunologi fokuserar på SADS-CoV N-proteinets roller vid IFN-undertryckning under infektion. I den här studien, forskare jämförde aminosyralikheterna mellan N -proteiner från olika CoV som tillhör fyra olika släkten. Målen för varje N -protein associerat med IFN -signalering diskuteras också. Mekanismen för IFN-inhibering har bestämts med användning av SADS-CoV N-protein via jämförande analys.
Forskarna i denna studie avslöjade att för att undertrycka IFN -signalering, steget för PAMP -igenkänning är ett kritiskt mål för N -proteinet. För detta ändamål, interaktionen mellan SADS-CoV N-proteinet och RIG-I utlöser ubiquitination, vilket främjar proteasomberoende nedbrytning. Detta leder till undertryckande av värdens IFN -svar.
Denna studie har också utvärderat flera N-proteiner av SADS-CoV för att bedöma deras förmåga att hämma IFN-svar. I sista hand, forskarna fann att hämningen av detta svar inte är beroende av aminosyrasekvenslikheten. Till exempel, det finns en 91,2% aminosyralikhet mellan SARS-CoV-2 och SARS-CoV. Dock, när det gäller mekanismerna för SARS-CoV-2, N -proteinet kan hämma aktiviteten hos IFN -promotorn, som annars induceras av RIG-I, MAVS, TBK1, och IKKϵ, medan SARS-CoV N-protein misslyckades med att göra det.
Ett sådant resultat belyser vikten av tertiär struktur för att definiera proteinfunktion. Det finns fortfarande ett gap i den aktuella forskningen i en fullständig förståelse av den tertiära strukturen för CoV N -proteinet. Även om detta kan vara sant, data om strukturer för N -terminaldomänen (NTD) och C -terminaldomän (CTD) för olika CoV N -proteiner är för närvarande tillgängliga.
Resultatet av den aktuella forskningen är i linje med tidigare rapporter. För detta ändamål, tidigare studier har visat att N -proteinet i PEDV provocerar värd -IFN -svar genom att interagera med TBK1 direkt. Dessutom, N-proteinet i SARS-CoV interagerar direkt eller indirekt med TRIM25 och proteinaktivatorn för proteinkinas R (PACT) för att aktivera RIG-I.
Den föreliggande forskningen har också föreslagit att N-proteinet i SADS-CoV riktar sig mot de första stegen i IFN-svaret och kan direkt störa aktiveringen av RIG-I. Denna jämförande analys har också visat att interferens med RIG-I kan vara den ledande metoden för N-proteinet i SADS-CoV för att undertrycka RIG-I-liknande receptorsignalering (RLR). Denna studie observerade att SADS-CoV N-proteinet riktar sig mot RIG-I för att hämma IFN-β-promotoraktivitet.
 Mikroplast identifierad i mänskligt avfall för första gången
Mikroplast identifierad i mänskligt avfall för första gången
 Använda FLUOstar Omega för att studera nya tarmbakterier som kan påverka vår hälsa
Använda FLUOstar Omega för att studera nya tarmbakterier som kan påverka vår hälsa
 Crohns sjukdom
Crohns sjukdom
 Tarmbakteriell profil kan förutsäga tarmskador efter strålbehandling
Tarmbakteriell profil kan förutsäga tarmskador efter strålbehandling
 Forskare utvecklar ett tillvägagångssätt för att vaccinera mot tarminflammation
Forskare utvecklar ett tillvägagångssätt för att vaccinera mot tarminflammation
 Forskare identifierar en bakterie med anti-SARS-CoV-2-aktivitet in vitro:Dolosigranulum pigrum
Forskare identifierar en bakterie med anti-SARS-CoV-2-aktivitet in vitro:Dolosigranulum pigrum
 Organchips -teknik förbättrar studier av tarmen för personlig medicin
Forskare har utvecklat tekniken för orgelchips med hjälp av donatorhärledda tarmceller som ger fördelar jämfört med organoider och ger nya möjligheter för personlig medicin. Inuti tarmen
Organchips -teknik förbättrar studier av tarmen för personlig medicin
Forskare har utvecklat tekniken för orgelchips med hjälp av donatorhärledda tarmceller som ger fördelar jämfört med organoider och ger nya möjligheter för personlig medicin. Inuti tarmen
 Studie tyder på koppling mellan probiotisk användning och "hjärntimmighet"
En studie gjord vid Medical College of Georgia vid Augusta University har visat att probiotiskt intag kan resultera i en betydande uppbyggnad av tunntarmbakterier som leder till dimmighet i hjärnan.
Studie tyder på koppling mellan probiotisk användning och "hjärntimmighet"
En studie gjord vid Medical College of Georgia vid Augusta University har visat att probiotiskt intag kan resultera i en betydande uppbyggnad av tunntarmbakterier som leder till dimmighet i hjärnan.
 E. coli superbug sprider sig genom dålig toaletthygien,
inte genom mat En ny studie publicerad i Lancet infektionssjukdomar den 22 oktober, 2019, säger att en vanlig superbug som orsakar mer än 5, 000 fall av matförgiftning i England varje år sprids inte
E. coli superbug sprider sig genom dålig toaletthygien,
inte genom mat En ny studie publicerad i Lancet infektionssjukdomar den 22 oktober, 2019, säger att en vanlig superbug som orsakar mer än 5, 000 fall av matförgiftning i England varje år sprids inte